The development of multi-cellular organisms is considered to be one of the key milestones in the path from the origin of life to the emergence of a technological civilisation able to spread life throughout the universe. Robin Hanson cited it as item 6 in his description of the “Great Filter” which must be passed before life can spread from its place of origin outward to the stars.
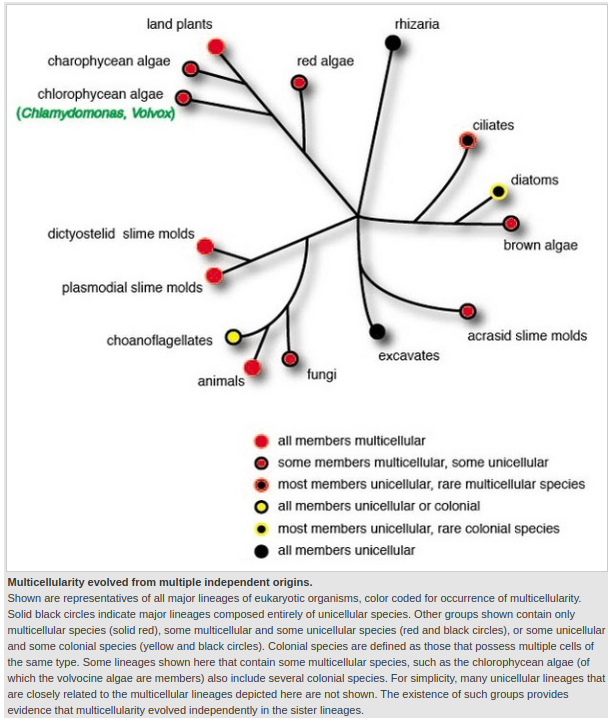
Unlike many of the events in the development of life which appear to have happened only once, with all organisms having that property descended from a common ancestor, multicellularity has evolved numerous times among a wide variety of evolutionary paths. The mechanisms leading to its appearance and the evolutionary adaptation of single cells to become parts of a complex organism have remained largely mysterious.
A paper published in the 2023-05-10 issue of Nature, “De novo evolution of macroscopic multicellularity”, reports an ongoing experiment called “Multicellularity Long-Term Evolution Experiment (MuLTEE)”, running since 2018 at the Georgia Institute of Technology (Georgia Tech), in which 3000 generations of snowflake yeast (Saccharomyces cerevisiae) were bred in an anaerobic environment with 600 rounds of selection for organism size. Here is the abstract:
While early multicellular lineages necessarily started out as relatively simple groups of cells, little is known about how they became Darwinian entities capable of sustained multicellular evolution. Here we investigate this with a multicellularity long-term evolution experiment, selecting for larger group size in the snowflake yeast (Saccharomyces cerevisiae) model system. Given the historical importance of oxygen limitation, our ongoing experiment consists of three metabolic treatments—anaerobic, obligately aerobic and mixotrophic yeast. After 600 rounds of selection, snowflake yeast in the anaerobic treatment group evolved to be macroscopic, becoming around 2\times 10^4 times larger (approximately mm scale) and about 10^4-fold more biophysically tough, while retaining a clonal multicellular life cycle. This occurred through biophysical adaptation—evolution of increasingly elongate cells that initially reduced the strain of cellular packing and then facilitated branch entanglements that enabled groups of cells to stay together even after many cellular bonds fracture. By contrast, snowflake yeast competing for low oxygen remained microscopic, evolving to be only around sixfold larger, underscoring the critical role of oxygen levels in the evolution of multicellular size. Together, this research provides unique insights into an ongoing evolutionary transition in individuality, showing how simple groups of cells overcome fundamental biophysical limitations through gradual, yet sustained, multicellular evolution.
The full text of the paper is behind a Springer paywall, but you can read the original submission to Nature for free at bioRχiv. A popular description of the experiment was released by Georgia Tech the day of the Nature publication, “A Journey to the Origins of Multicellular Life: Long-Term Experimental Evolution in the Lab”.
This type of biophysical evolution is a pre-requisite for the kind of large multicellular life that can be seen with the naked eye. Their study is the first major report on the ongoing Multicellularity Long-Term Evolution Experiment (MuLTEE), which the team hopes to run for decades.
“Conceptually, what we want to understand is how simple groups of cells evolve into organisms, with specialization, coordinated growth, emergent multicellular behaviors, and life cycles – the stuff that differentiates a pile of pond scum from an organism that is capable of sustained evolution,” [co-author William C.] Ratcliff said. “Understanding that process is a major goal of our field.”
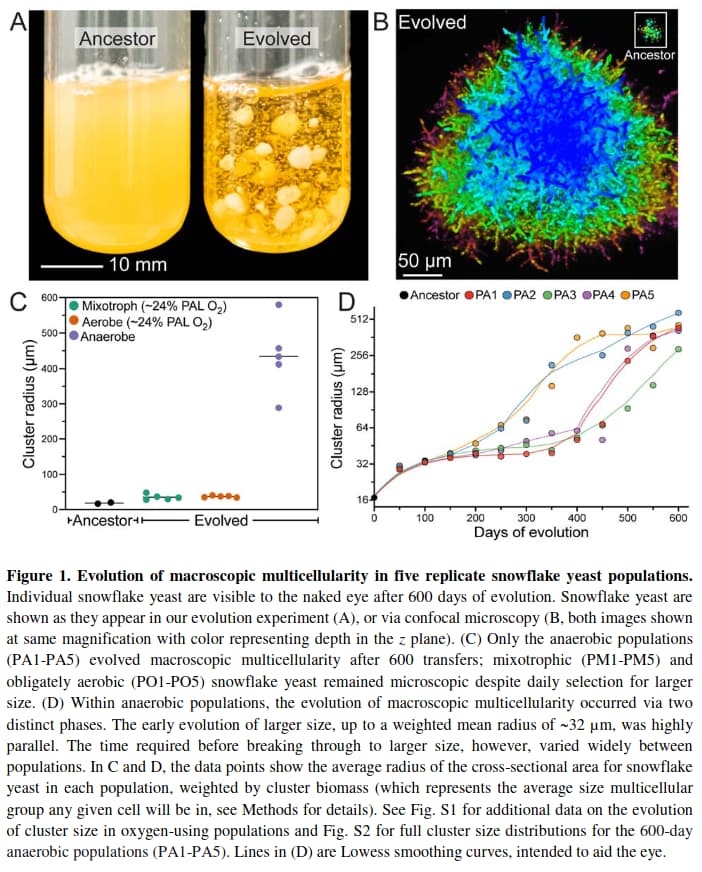
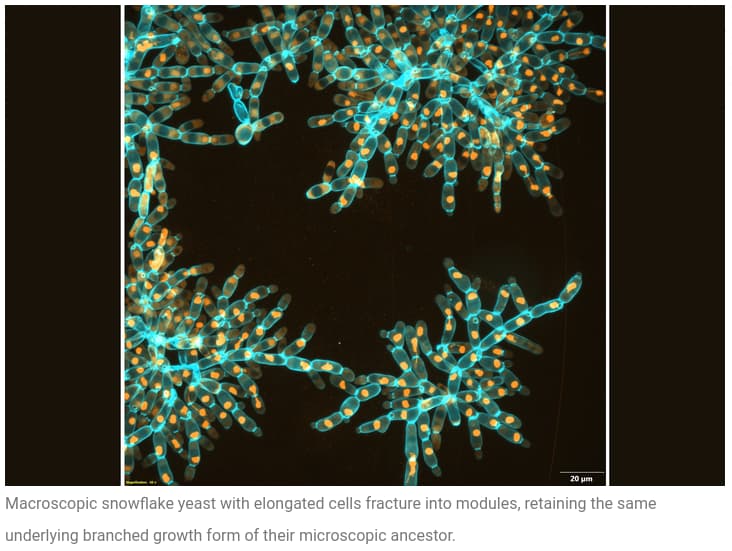
It is sometimes claimed that evolution only refines what is already present in an organism, but is incapable of invention and innovation. Yet here, in a laboratory experiment running only five years, a single-cell organism has modified itself to live in colonies, becoming 20,000 times larger and 10,000 times tougher than the organism in the wild, changing its form into an elongated shape with the ability to entangle to hold colonies together. Just image what you could do with a planet-sized laboratory and a few tens of millions of years.